Клетками хронического воспаления являются

Клетки участвующие в хроническом воспалении — макрофаги и другие клетки
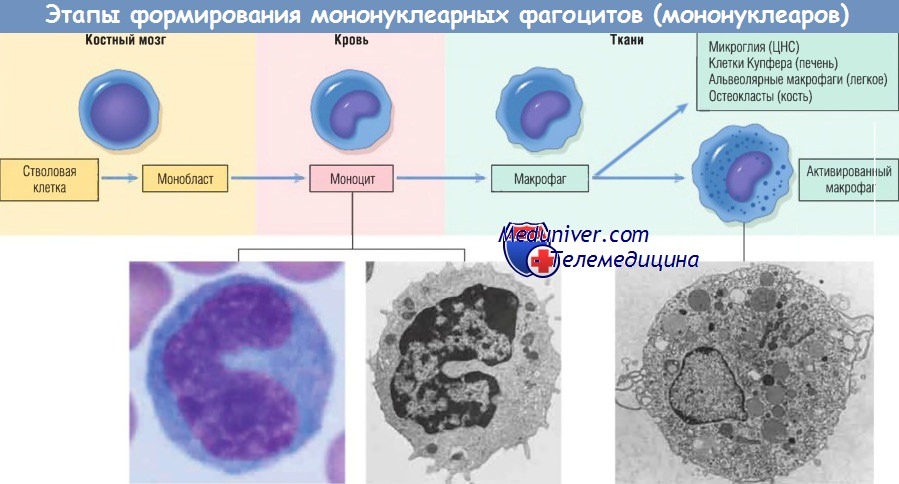
Макрофаг — главная клетка, участвующая в процессе хронического воспаления. Макрофаги являются частью системы мононуклеарных фагоцитов (иногда называемой ретикулоэндотелиальной системой), состоящей из клеток, близкородственных клеткам костного мозга, включая моноциты и тканевые макрофаги.
Тканевые макрофаги диффузно разбросаны по соединительной ткани или расположены в таких органах, как печень (клетки Купфера), селезенка и лимфатические узлы (гистиоциты синусов), легкие (альвеолярные макрофаги), а также в ЦНС (микроглия). Мононуклеарные фагоциты происходят из клеток-предшественников костного мозга, которые дифференцируются в моноциты. Из крови моноциты мигрируют в разные ткани и дифференцируются в макрофаги.
Период полужизни моноцитов длится около 1 дня, когда как период жизни тканевых макрофагов составляет несколько месяцев или лет. Превращение стволовой клетки костного мозга в тканевый макрофаг регулируется различными факторами роста и дифференцировки, цитокинами, молекулами адгезии и клеточными взаимодействиями.
Моноциты начинают мигрировать во внесосудистые ткани на ранних стадиях развития острого воспаления и в течение 48 час могут стать доминирующим типом клеток инфильтрата. Экстравазация моноцитов управляется теми же стимулами, которые участвуют в миграции нейтрофилов и представлены молекулами адгезии и химическими медиаторами с хемотаксическими и активирующими свойствами.
При попадании моноцита во внесосудистую ткань он подвергается трансформации в большую фагоцитарную клетку — макрофаг. Макрофаги активируются различными стимулами, включая микробные продукты, активирующие TLR и другие клеточные рецепторы, цитокины (например, IFN-y), секретированные сенсибилизированными Т-лимфоцитами и естественными клетками-киллерами, и другие химические медиаторы.
В результате активации макрофагов происходят элиминация таких повреждающих агентов, как микробы, и инициация процессов репарации, а также повреждение ткани (при хроническом воспалении).
Активация макрофагов приводит также к повышению уровней лизосомных ферментов и активных форм кислорода и азота, продукции цитокинов, факторов роста и других медиаторов воспаления. Некоторые из этих продуктов токсичны для микробов и организма (например, активные формы кислорода и азота) или ВКМ (протеазы); другие вызывают миграцию клеток (например, цитокины, хемотаксический стимул); третьи влияют на пролиферацию фибробластов, отложение коллагена и ангиогенез (например, факторы роста).
Разные популяции макрофагов выполняют различные функции: некоторые необходимы при киллинге микробов и воспалении, другие очень важны при репарации.
Впечатляющий арсенал медиаторов делает макрофаги мощным союзником организма в защите от непрошеных гостей, но в то же время при неправильной активации макрофаги могут вызывать значительное разрушение ткани, которое является одним из признаков хронического воспаления. Процесс разрушения ткани сам по себе может активировать каскад воспалительной реакции, поэтому одновременно могут присутствовать признаки и острого, и хронического воспаления.
При недолгом процессе воспаления, если стимул удален, макрофаги в итоге либо умирают, либо уходят в лимфатические сосуды и лимфоузлы. При хроническом воспалении происходит накопление макрофагов в результате их мобилизации из кровотока и местной пролиферации в очагах воспаления.
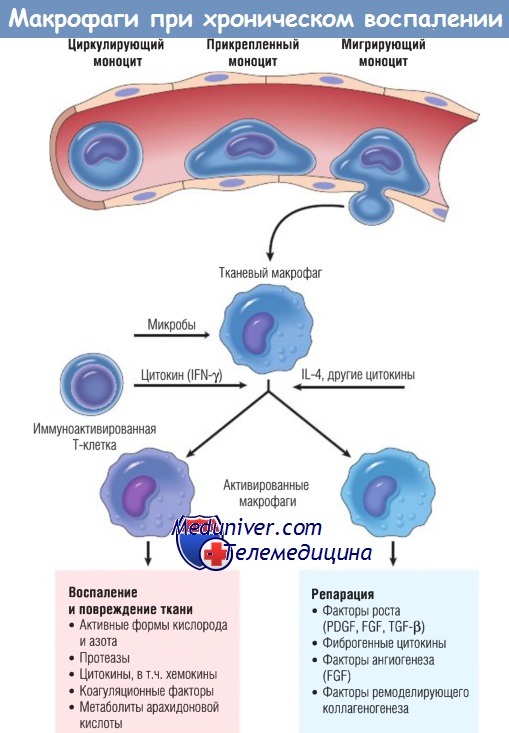
Роль активированных макрофагов при хроническом воспалении.
Макрофаги активируются неиммунными стимулами, например эндотоксином, или цитокинами из иммуноактивированных Т-клеток (особенно IFN-y).
Перечислена продукция активированных макрофагов.
FGF — фактор роста фибробластов; IFN — интерферон; IL — интерлейкин;
PDGF — тромбоцитарный фактор роста; TGF — трансформирующий фактор роста.
В развитии хронического воспаления участвуют и другие типы клеток: лимфоциты, клетки плазмы крови, эозинофилы и тучные клетки:
— лимфоциты мобилизуются в ходе как антителоопосредованных, так и клеточно-опосредованных иммунных реакций. Антиген-стимулированные лимфоциты (эффекторные клетки и клетки памяти) различных типов (Т- и В-клетки) используют разные пары адгезивных молекул (селектинов, интегринов и их лигандов) и хемокины для миграции в очаг воспаления. Цитокины активированных макрофагов, в основном TNF, IL-1 и хемокины, усиливают мобилизацию лейкоцитов, запуская стадию персистенции воспалительной реакции.
Лимфоциты и макрофаги взаимодействуют по реверсивному типу, что играет важную роль в развитии хронического воспаления. Макрофаги презентируют антигены Т-клеткам и продуцируют мембранные молекулы (костимуляторы) и цитокины (например, IL-12), которые стимулируют ответ Т-клеток. Активированные Т-клетки продуцируют цитокины, некоторые из которых мобилизуют моноциты из кровотока, и IFN-y — мощный активатор макрофагов. В результате этих взаимодействий Т-клеток и макрофагов в воспалительную реакцию вовлекается иммунная система, что делает реакцию хронической и тяжелой. Термин «иммунное воспаление» отражает наличие выраженного иммунного компонента в воспалительном процессе, например ответа Т- и В-клеток;
— клетки плазмы крови образуются из активированных В-клеток и продуцируют антитела, направленные либо против персистирующих чужеродных или собственных антигенов в очаге воспаления, либо против компонентов поврежденной ткани. При некоторых сильных воспалительных реакциях скопление лимфоцитов, антигенпрезентирующих клеток и клеток плазмы крови может напоминать морфологическое строение лимфоидного органа, особенно лимфоузла, содержащего хорошо сформированные герминативные центры.
Подобные очаги скопления лимфоцитов называют третичными лимфоидными органами. Такой тип лимфоидного органогенеза часто встречается в синовиальных оболочках у пациентов, длительно страдающих ревматоидным артритом;
— эозинофилами изобилуют иммунные реакции, опосредованные IgE и паразитарными инфекциями. Особо важный хемокин для мобилизации эозинофилов — эотаксин. Эозинофилы содержат гранулы, имеющие главный основной белок (высокоактивный катионный белок), который токсичен для паразитов, но может вызвать лизис эпителиальных клеток млекопитающих. Таким образом, эозинофилы контролируют паразитарные инфекции, но также участвуют в повреждении ткани при иммунных реакциях, например аллергии;
— тучные клетки широко распространены в соединительных тканях по всему телу и могут участвовать как в острых, так и в хронических воспалительных реакциях. Тучные клетки экспрессируют на своей поверхности рецептор FceRI, связывающийся с Fc-фракцией антитела IgE. При реакции гиперчувствительности немедленного типа антитела IgE связываются с клеточными Fc-peцепторами, специфично распознающими антиген, клетка дегранулируется и высвобождает медиаторы — гистамин и простагландины. Этот тип ответа наблюдается при пищевой аллергии, укусах насекомых, лекарственной аллергии, иногда с ужасающими исходами (например, анафилактическим шоком).
Тучные клетки тоже присутствуют при реакциях хронического воспаления и секретируют множество цитокинов, т.е. они способны как усиливать, так и ограничивать воспалительную реакцию в разных ситуациях.
Как правило, присутствие нейтрофилов характерно для острого воспаления, но и при многих формах хронического воспаления, длящегося месяцами, выявляется большое количество нейтрофилов, индуцированных либо персистирующими микробами, либо медиаторами, продуцированными активированными макрофагами и Т-лимфоцитами. При хронических бактериальных инфекциях костей (остеомиелите) нейтрофильный экссудат может сохраняться многие месяцы. Нейтрофилы также важны при хроническом воспалении в легких, индуцированном курением или другим стимулом.
Помимо мононуклеарного характера клеточного инфильтрата при хроническом воспалении часто происходит пролиферация кровеносных и лимфатических сосудов. Этот процесс стимулируют факторы роста, продуцируемые макрофагами и эндотелиальными клетками.
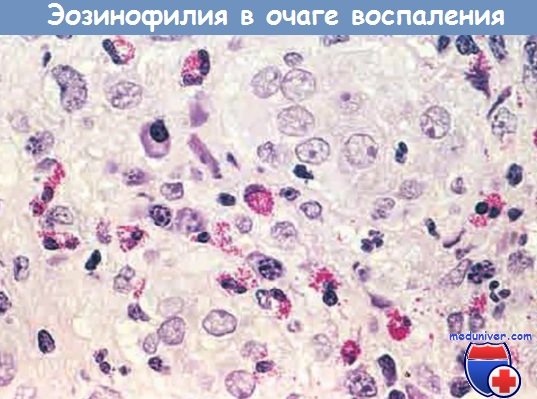
Очаг воспаления с большим количеством эозинофилов (эозинофилия)
Взаимодействия макрофагов и лимфоцитов при хроническом воспалении.
Активированные Т-клетки продуцируют цитокины, которые мобилизуют макрофаги (TNF, IL-17, хемокины), и IFN-y, который является мощным активатором макрофагов.
Разные типы Т-клеток (Тh1 и Тh17) могут продуцировать различные типы цитокинов.
В свою очередь, активированные макрофаги стимулируют Т-клетки, презентируя им антигены через цитокины (например, IL-12).
IFN — интерферон; IL — интерлейкин; TNF — фактор некроза опухоли.
— Рекомендуем ознакомиться со следующей статьей «Причины и механизмы гранулематозного воспаления. Что такое гранулема?»
Оглавление темы «Патофизиология»:
- Клетки участвующие в хроническом воспалении — макрофаги и другие клетки
- Причины и механизмы гранулематозного воспаления. Что такое гранулема?
- Влияние воспаления на организм в целом (системные симптомы воспаления)
- Варианты воспалительной реакции и ее последствия
- Пути заживления и обновления тканей — репарация, регенерация
- Регуляция деления и пролиферации клеток в тканях
- Виды стволовых клеток и их характеристика
- Эмбриональные стволовые клетки и их характеристика
- Индуцированные плюрипотентные стволовые клетки (ИПСК) и их характеристика
- Где присутствуют стволовые клетки у взрослых?
Бывают случаи, когда в воспалительных инфильтратах с самого начала скапливаются не полиморфно-ядерные лейкоциты, а моноциты, лимфоциты и их производные. Формирование такого скопления мононуклеарных клеток, получившего название «гранулема», является предпосылкой к долгому течению воспаления. Хроническое воспаление служит иллюстрацией справедливости высказывания И.И. Мечникова: «воспаление — реакция защитная по своей биологической сути, но, к сожалению, для организма не всегда достигающая совершенства».
В отличие от острого воспаления хроническое воспаление начинаетсяне с нарушений микроциркуляции и описанных ранее событий в сосудистом русле, а со скопления критического числараздраженных (активированных) макрофаговв одном месте.
Стойкое раздражение макрофагов можно вызвать разными способами.
• Ряд микробов поглощается макрофагами, но, оказавшись в их фагосомах, не погибает и получает возможность длительно персистировать и размножаться внутри клетки (это возбудители туберкулеза, проказы, листериоза, токсоплазмоза и многие другие). Макрофаги, содержащие микробы, переходят в активное состояние и начинают секретировать медиаторы воспаления.
• Макрофаги могут поглощать неинфекционные частицы, которые клетка не в состоянии расщепить или выбросить в среду (сложные полисахаридные комплексы — коррагенан из морских водорослей, декстран, зимозан из пекарских дрожжей). После внутривенного введения мышам гранул зимозана они захватываются макрофагами-резидентами (клетками Купфера) печени и макрофагами интерстиция легкого и активируют их. Через 2-3 суток вокруг таких макрофагов, как вокруг эпицентров, начинают скапливаться попавшие с кровью моноциты и формируется то, что принято называть гранулемой, или мононуклеарным инфильтратом. Привлечение новых моноцитов/макрофагов в зону локализации активированных макрофагов связано с веществами, вызывающими хемотаксис. Они выделяются активными макрофагами в готовом виде (LTC4, LTD4, PGE2) либо в виде предшественников: С2, С4, С5, С6 компоненты комплемента, которые превращаются в СЗа, С5а, С567-фракции с высокой хемотаксической активностью под действием протеаз, секретируемых теми же макрофагами.
Лизосомальные ферменты, секретируемые макрофагами, как коллагеназа, расщепляют коллаген. Продукты частичной деградации коллагена обладают мощной способностью притягивать свежие моноциты в очаг воспаления.
Активированные макрофаги секретируют биоокислители, которые запускают перекисное окисление липидов в мембранах других клеток в зоне инфильтрации. Однако простое повышение хемотаксиновв каком-то участке ткани еще не означало бы притока туда новых клеток-эффекторов воспаления из крови. Нужно, чтобы наряду с формированием градиента этих веществ произошло повышение проницаемостимикрососудов, из которых мононуклеарные лейкоциты могли бы поступать в зону локализации раздраженных макрофагов. Активированные макрофаги повышают проницаемость микрососудов, продуцируя LTC4, LTD4, фактор агрегации тромбоцитов, О2*-, коллагеназу и активатор плазминогена, разрыхляющие капилляросоединительнотканный барьер. Они либо разуплотняют базальную мембрану капилляров, либо сокращают эндотелиальные клетки и обнажают межэндотелиальные щели, либо действуют тем и другим способом. В результате облегчается выход лейкоцитов из крови и их передвижение в область высокой концентрации хемотаксинов, где они присоединяются к другим клеткам инфильтрата. Моноциты, придя в инфильтрат, выделяют
фибронектин. Благодаря этому они прочно связываются с матриксом соединительной ткани, прежде всего с коллагеновыми волокнами. Они как бы «становятся на якорь». В английской литературе такое обездвиживание клеток даже получило название «заякоривание»(от англ. anchor — якорь). Это очень важный момент, ибо «на ходу» фагоциты «не успевают решить тех проблем», которые возникают перед ними в очаге воспаления.
Фагоцитоз протекает наиболее эффективно лишь после того, как моноциты закрепляются и распластываются на структурах соединительной ткани. Таким образом, активные макрофаги не только запускают, но и детерминируют весь процесс хронического воспаления. Однако в реальных условиях макрофаги работают не в изоляции, а в комплексе с другими типами клеток, входящих в состав воспалительного инфильтрата (гранулемы) (рис. 10-16, см. цв. вклейку).
Лучше всего изучена функциональная кооперация между макрофагамии лимфоцитами:
1. Прежде всего эти клетки вступают в тесное взаимодействие в специфическом иммунном ответе, развивающемся при инфекционном воспалении. Макрофаги поглощают и частично разрушают микробные антигены в своих фаголизосомах. В модифицированном виде эти антигены вновь всплывают на цитоплазматическую мембрану макрофага, где они вступают в комплексную связь с особыми белками. Только в таком сочетании антиген распознается Т-лимфоцитами. Это взаимодействие макрофага и Т-лимфоцитов в очаге хронического воспаления можно назвать антигензависимым. Оно проявляет себя наиболее зримо при тех формах хронического воспаления, которые возникают при микробном заражении и протекают с явлениями гиперчувствительности замедленного типа (ГЗТ).
2. Наряду с этим макрофаги связаны с лимфоцитами не только через антигены, но и через свои секреты. Макрофаги выделяют вещества (например, IL-1), усиливающие рост лимфоцитов и повышающие их активность.
3. В то же время активно пролиферирующие лимфоциты выделяют лимфокины, которые активируют макрофаги и резко усиливают их эффекторные функции в очаге хронического воспаления:
• фактор торможения миграции макрофагов повышает адгезивность мембран макрофагов и дает им возможность прочно за-
цепиться за субстрат. Тот же фактор растормаживает секрецию макрофагами медиаторов воспаления;
• фактор, усиливающий агрегацию макрофагов, их пролиферацию, слияние макрофагов друг с другом с образованием гигантских многоядерных клеток, столь характерных для очагов хронического воспаления. В частности, таких клеток особенно много в туберкулезных инфильтратах в легких;
• лимфокины повышают микробицидный потенциал макрофагов, и клетки начинают убивать микробы, которые до того в них безнаказанно паразитировали. Это связано с тем, что, вопервых, лимфокины усиливают слияние фагосом с лизосомами (имеющиеся в фагосомах живые микробы такого слияния часто не выдерживают и погибают). Во-вторых, лимфокины повышают активность НАДФН-оксидазы, которая отвечает за образование в мембранах фагосом О2*- и Н2О2 — основных микробицидных факторов фагоцитов.
Пути запуска и развития острого и хронического воспаленияпринципиально отличаются:
1. При остром воспалении процесс запускается «от сосудов», тогда как при хроническом воспалении — с территории соединительной ткани, где находятся активные макрофаги.
2. Ведущей клеткой острого воспаления — эффектором — является нейтрофил, а хронического воспаления — активный макрофаг. Все другие клетки мезенхимы (тучные, лимфоциты, эозинофилы) тоже вносят свой вклад в реализацию процесса, модулируя реактивность нейтрофилов и макрофагов.
3. Острое воспаление заканчивается быстро, в считанные дни, если не возникнет осложнений в виде гнойной полости (абсцесса).
4. Хроническое воспаление не может закончиться быстро по следующим причинам:
• во-первых, макрофаги в очаге воспаления имеют длительный жизненный цикл, который исчисляется неделями, месяцами и даже годами. Вначале, на этапе зарождения, в гранулему приходят свежие моноциты с кровью, лимфоциты — с кровью и лимфой. Они еще не обладают достаточно высокой микробицидной активностью. Затем гранулема постепенно зреет, и в ней накапливаются дифференцированные макрофаги, активно поглощающие микробы. Наконец, на заключительном этапе, в застарелой гранулеме число активно фагоцитирующих клеток уменьшается, но зато возрастает процент относительно
• инертных в смысле фагоцитоза эпителиоидных и гигантских многоядерных клеток; во-вторых, любая гранулема — это не «застывшее» образование. В нее постоянно следуют потоком все новые и новые моноциты с кровью из костного мозга. Если в гранулеме много активированных макрофагов, приток будет превышать отток клеток из гранулемы. Дело в том, что раздраженные макрофаги усиленно вырабатывают особые гемопоэтины. Они стимулируют образование фагоцитов в костном мозгу. К их числу относится колониестимулирующий фактор Меткалфа. Поэтому пока раздраженные макрофаги «работают», баланс будет смещен в сторону притока клеток в инфильтрат, и его рассасывание невозможно. Если макрофаги выделяют много биоокислителей в среду своего обитания, они могут не только санировать очаг, но и повредить собственные клетки организма. При гиперпродукции Н2О2 и О2*- эти факторы могут ускользать из фагосом в цитозоль макрофага и вести к его гибели. В целях предупреждения такой ситуации в макрофагах имеется система аварийной нейтрализации избытка биоокислителей. В нее входят ферменты: каталаза, глутатионпероксидаза и глутатионредуктаза. В частности, под действием глутатионредуктазы осуществляется нейтрализация перекиси водорода в реакции 2 ГН + Н2О2 — Г-Г + 2Н2О, где Г — глутатион. Фермент супероксиддисмутаза нейтрализует супероксидный анион-радикал (О2*-) в реакции О2*- + О2*-+ 2Н+ — Н2О2 + О2. Когда системы антиоксидантной защиты не срабатывают, это ведет к персистенции воспаления.
Хроническое воспаление может продолжаться в течение всей жизни. Периодически оно обостряется, когда в очаг поступают нейтрофилы и свежие макрофаги с высокой провоспалительной активностью. В очаге мононуклеарной инфильтрации идет деструкция соединительной ткани. В ответ на это происходит разрастание волокнистых структур. В конечном счете может развиться склероз с частичным или полным выключением специализированных функций органа. Этому способствует накопление в гранулеме особого класса макрофагов, секретирующих фибробластстимулирующие факторы. С такой ситуацией врачам приходится встречаться при циррозах печени после вирусных гепатитов, хронических пневмониях, хронических гломерулонефритах и других хронически протекающих воспалительных заболеваниях.